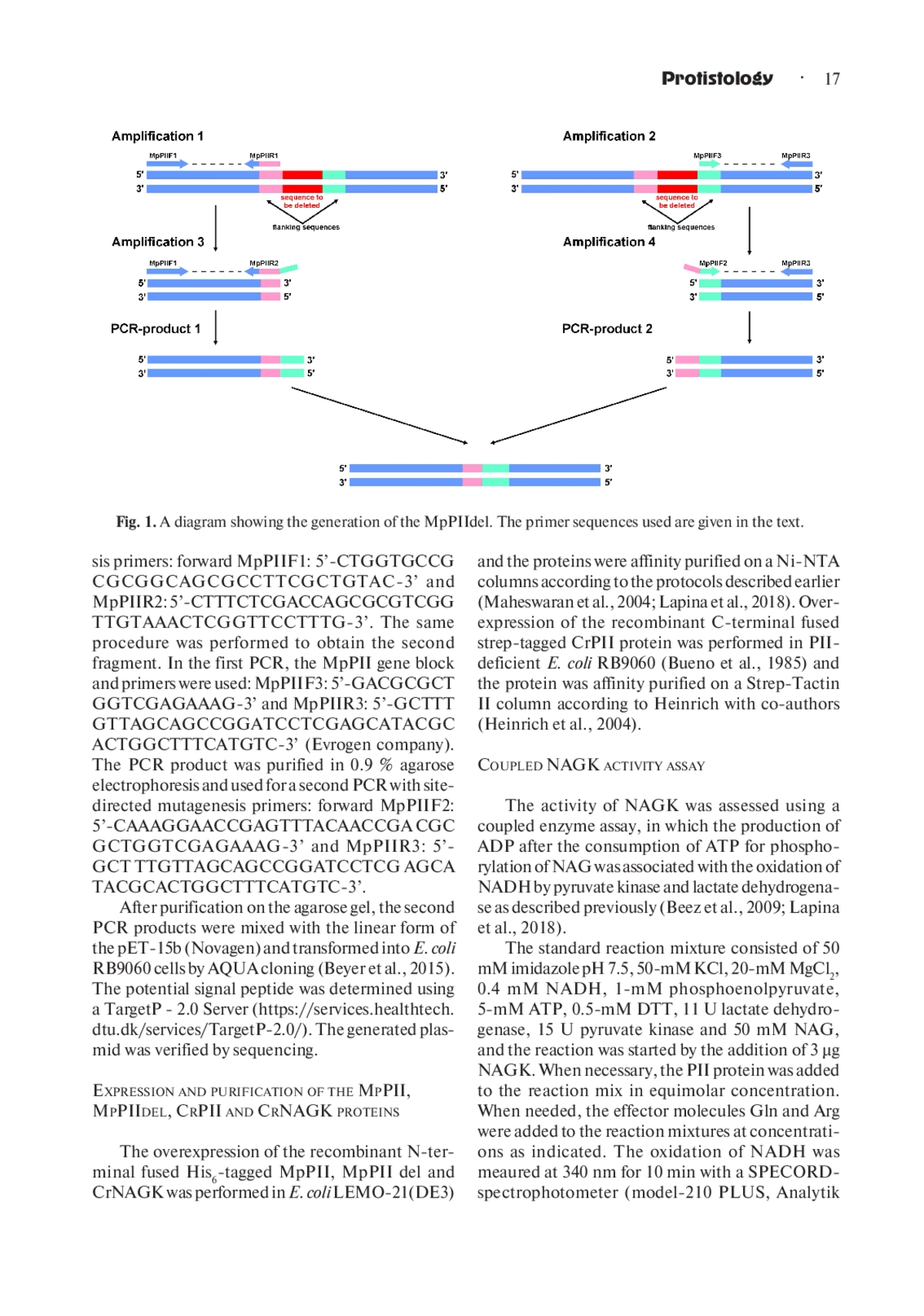
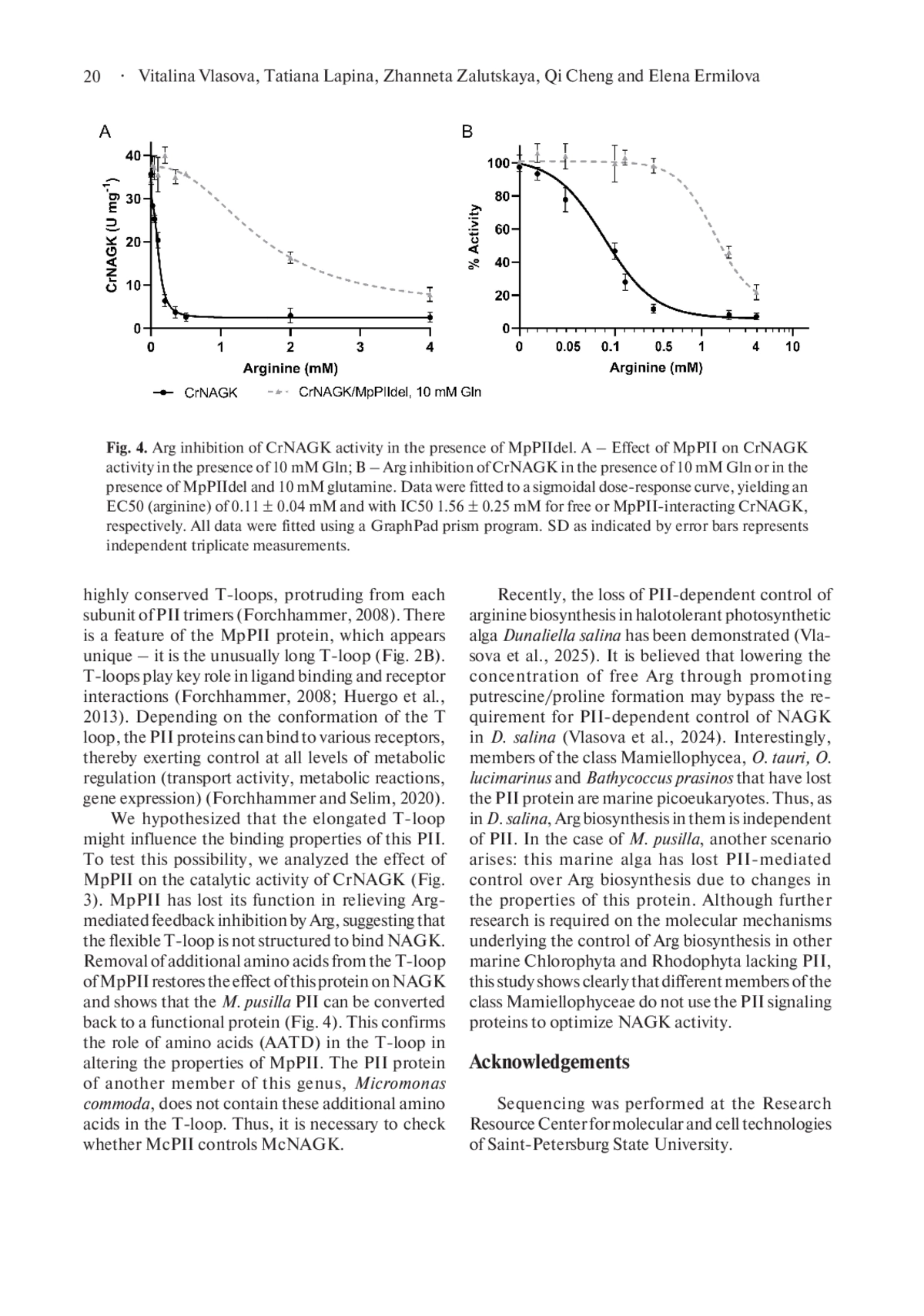
1. Beez S., Fokina O., Herrmann C. and Forchhammer K. 2009. N-acetyl-L-glutamate kinase (NAGK) from oxygenic phototrophs: PII signal transduction across domains of life reveals novel insights in NAGK control. J. Mol. Biol. 389 (4): 748- 758. DOI: 10.1016/j.jmb.2009.04.053
2. Beyer H.M., Gonschorek P., Samodelov S.L., Meier M. et al. 2015. AQUA Cloning: A versatile and simple enzyme-free cloning approach. PLoS One. 10 (9): e0137652. DOI: 10.1371/journal.pone.0137652
3. Blanc-Mathieu R., Verhelst B., Derelle E., Rombauts S. et al. 2014. An improved genome of the model marine alga Ostreococcus tauri unfolds by assessing Illumina de novo assemblies. BMC Genomics. 15 (1):1103. DOI: 10.1186/1471-2164-15-1103
4. Bueno R., Pahel G. and Magasanik B. 1985. Role of glnB and glnD gene products in regulation of the glnALG operon of Escherichia coli. J. Bacteriol. 164 (2): 816-822. DOI: 10.1128/jb.164.2.816-822.1985
5. Chellamuthu V.R., Alva V. and Forchhammer K. 2013. From cyanobacteria to plants: Conservation of PII functions during plastid evolution. Planta. 237 (2): 451-462. DOI: 10.1007/s00425-012-1801-0
6. Chellamuthu V.R., Ermilova E., Lapina T., Lüddecke J. et al. 2014. A widespread glutamine-sensing mechanism in the plant kingdom. Cell. 159 (5): 1188-1199. DOI: 10.1016/j.cell.2014.10.015
7. Chen Y.M., Ferrar T.S., Lohmeier-Vogel E.M., Morrice N. et al. 2006. The PII signal transduction protein of Arabidopsis thaliana forms an arginine regulated complex with plastid N-acetyl glutamate kinase. J. Biol. Chem. 281 (9): 5726-5733. DOI: 10.1074/jbc.M510945200
8. Fokina O., Chellamuthu V.R., Forchhammer K. and Zeth K. 2010. Mechanism of 2-oxoglutarate signaling by the Synechococcus elongatus PII signal transduction protein. Proc. Natl. Acad. Sci. USA. 107 (46): 19760-19765. DOI: 10.1073/pnas.1007653107
9. Forcada-Nadal A., Llácer J.L., Contreras A., Marco-Marín C. et al. 2018. The PII-NAGK- PipX-NtcA regulatory axis of Cyanobacteria: A tale of changing partners, allosteric effectors and non-covalent interactions. Front. Mol. Biosci. 5:91. DOI: 10.3389/fmolb.2018.00091
10. Forchhammer K. 2008. PII signal transducers: novel functional and structural insights. Trends Microbiol. 16 (2): 65-72. DOI: 10.1016/j.tim.2007.11.004
11. Forchhammer K. and Selim K.A. 2020. Carbon/ nitrogen homeostasis control in cyanobacteria. FEMS Microbiol. Rev. 44 (1): 33-53. DOI: 10.1093/femsre/fuz025
12. Guo X., Pang M., Zheng X. and Huang L. 2024. Micromonas, a small pigmented flagellate, predominates the nanoflagellate and photosynthetic picoeukaryote communities in the northern South China Sea. Environ. Microbiol. Rep. 16 (2): e13244. DOI: 10.1111/1758-2229.13244
13. Heinrich A., Maheswaran M., Ruppert U. and Forchhammer K. 2004. The Synechococcus elongatus PII signal transduction protein controls arginine synthesis by complex formation with N- acetylL-glutamate kinase. Mol. Microbiol. 52 (5): 1303-1314. DOI: 10.1111/j.1365-2958.2004.04058.x
14. Huergo L.F., Chandra G. and Merrick M. 2013. P(II) signal transduction proteins: nitrogen regulation and beyond. FEMS Microbiol. Rev. 37 (2): 251-83. DOI: 10.1111/j.1574-6976.2012.00351.x
15. Lapina T., Selim K.A., Forchhammer K. and Ermilova E. 2018. The PII signaling protein from red algae represents an evolutionary link between cyanobacterial and Chloroplastida PII proteins. Sci. Rep. 8 (1): 790. DOI: 10.1038/s41598-017-19046-7
16. Leliaert F., Verbruggen H. and Zechman F.W. 2011.Into the deep: New discoveries at the base of the green plant phylogeny. Bioessays. 33 (9): 683-692. DOI: 10.1002/bies.201100035
17. Llácer J.L., Contreras A., Forchhammer K., Marco-Marín C. et al. 2007. The crystal structure of the complex of PII and acetylglutamate kinase reveals how PII controls the storage of nitrogen as arginine. Proc. Natl. Acad. Sci. USA. 104 (45): 17644-17649. DOI: 10.1073/pnas.0705987104
18. Llácer J.L., Fita I. and Rubio V. 2008. Arginine and nitrogen storage. Curr. Opin. Struct. Biol. 18 (6): 673-681. DOI: 10.1016/j.sbi.2008.11.002
19. Maheswaran M., Urbanke C. and Forchhammer K. 2004.Complex formation and catalytic activation by the PII signaling protein of N-acetyl-L-glutamate kinase from Synechococcus elongatus strain PCC 7942. J. Biol. Chem. 279 (53): 55202-55210. DOI: 10.1074/jbc.M410971200
20. Marin B. and Melkonian M. 2010. Molecular phylogeny and classification of the Mamiellophyceae class. nov. (Chlorophyta) based on sequence comparisons of the nuclear- and plastid-encoded rRNA operons. Protist. 161 (2): 304-336. DOI: 10.1016/j.protis.2009.10.002
21. Mizuno Y., Moorhead G.B. and Ng K.K. 2007. Structural basis for the regulation of N-acetyl-glutamate kinase by PII in Arabidopsis thaliana. J. Biol. Chem. 282 (49): 35733-35740. DOI: 10.1074/jbc.M707127200
22. Moreau H., Verhelst B., Couloux A., Derelle E. et al. 2012. Gene functionalities and genome structure in Bathycoccus prasinos reflect cellular specializations at the base of the green lineage. Genome Biology. 13. R74. DOI: 10.1186/gb-2012-13-8-r74
23. Sant’Anna F.H., Trentini D.B., Weber S.S., Cecagno R. et al. 2009. The PII superfamily revised: a novel group and evolutionary insights. J. Mol. Evol. 68 (4): 322-336. DOI: 10.1007/s00239-009-9209-6
24. Selim K.A., Ermilova E. and Forchhammer K. 2020a. From cyanobacteria to Archaeplastida: New evolutionary insights into PII signalling in the plant kingdom. New Phytol. 227 (3): 722-731. DOI: 10.1111/nph.16492
25. Selim K.A., Lapina T., Forchhammer K. and Ermilova E. 2020b.Interaction of N-acetyl-l-glutamate kinase with the PII signal transducer in the non-photosynthetic alga Polytomella parva: co-evolution towards a hetero-oligomeric enzyme. FEBS J. 287 (3): 465-482. DOI: 10.1111/febs.14989
26. Sugiyama K., Hayakawa T., Kudo T., Ito T. and Yamaya T. 2004.Interaction of N-acetylglutamate kinase with a PII-like protein in rice. Plant Cell Physiol. 45 (12): 1768-1778. DOI: 10.1093/pcp/pch199
27. Truan D., Huergo L.F., Chubatsu L.S., Merrick M. et al. 2010. A new PII protein structure identifies the 2-oxoglutarate binding site. J. Mol. Biol. 400 (3): 531-539. DOI: 10.1016/j.jmb.2010.05.036
28. Vlasova V., Lapina T., Cheng Q. and Ermilova E. 2025. Loss of PII-dependent control of arginine biosynthesis in Dunaliella salina. Plant Sci. 351. 112327. DOI: 10.1016/j.plantsci.2024.112327
29. Vlasova V., Lapina T., Zalutskaya Z., Puzanskiy R. et al. 2024. Control of arginine biosynthesis in the green alga Dunaliella salina. Protistology. 18 (3): 212-220. DOI: 10.21685/1680-0826-2024-18-3-3
30. Worden A.Z., Lee J.H., Mock T., Rouzé P. et al. 2009. Green evolution and dynamic adaptations revealed by genomes of the marine picoeukaryotes Micromonas. Science. 324 (5924): 268-272. DOI: 10.1126/science.1167222
31. Zeth K., Fokina O. and Forchhammer K. 2014. Structural basis and Target-Specific modulation of ADP Sensing by the Synechococcus elongatus PII signaling protein. J. Biol. Chem. 289: 8960-72.