

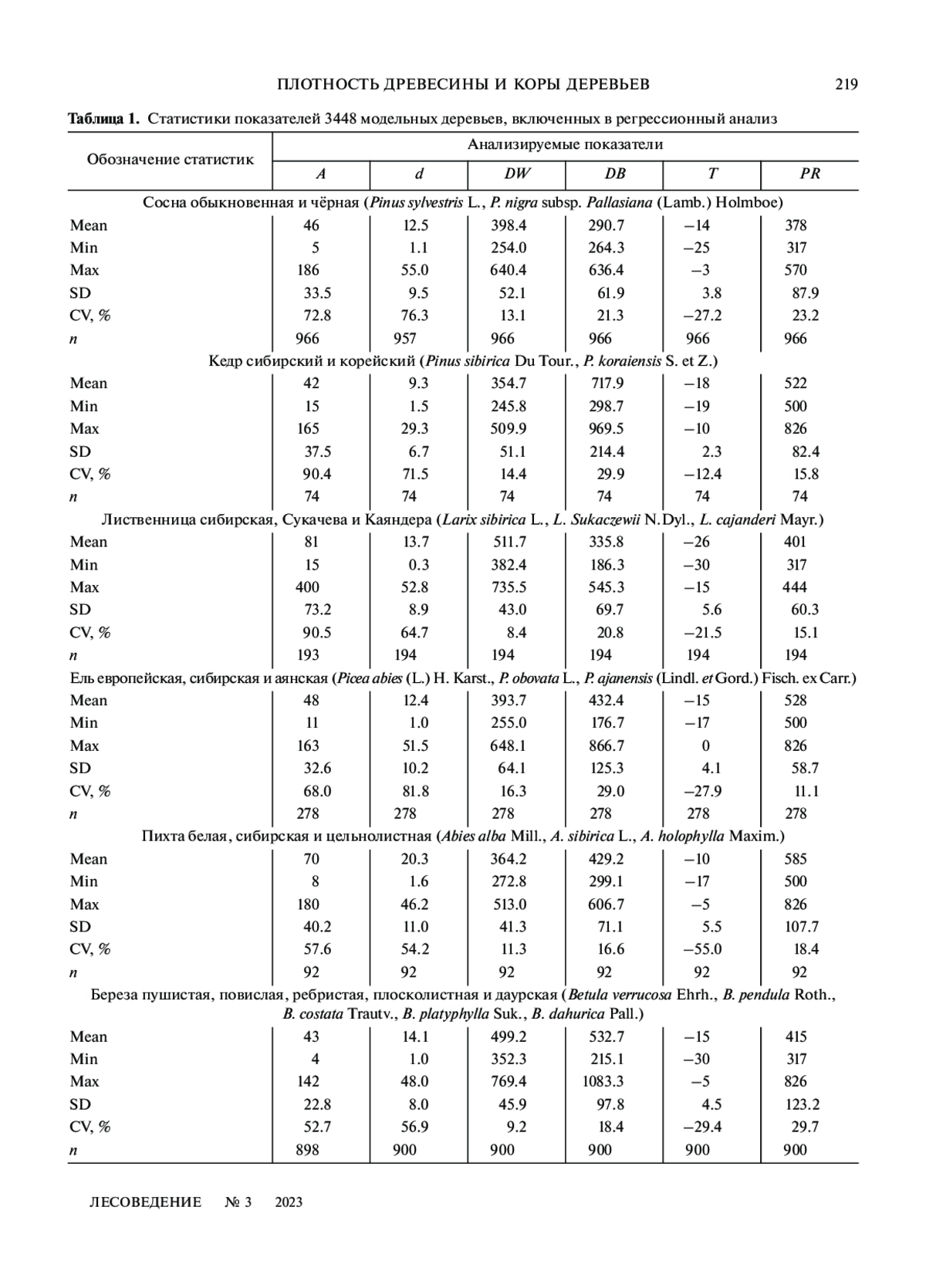
В условиях изменяющегося климата возрастает биосферная роль лесного покрова и актуальность исследований углерододепонирующей способности мировых лесов. Эти исследования содержат оценку биологической продуктивности деревьев и древостоев, включающей не только фитомассу, но и базисную плотность (БП) древесины и коры стволов. В нашем исследовании разработаны аллометрические модели БП древесины и коры деревьев 9 лесообразующих древесных видов Северной Евразии, имеющие такие независимые переменные, как возраст дерева, диаметр ствола, а также среднюю температуру января и среднегодовые осадки. Применена структура модели смешанного типа, в которой принадлежность исходных данных к каждому из древесных видов кодируется набором фиктивных переменных. На основе принципа пространственно-временного замещения полученные закономерности изменения БП в пространственных климатических градиентах использованы для прогноза их изменения в темпоральных градиентах. Подтверждено действие закона лимитирующего фактора Либиха при прогнозировании БП в пространственных и темпоральных климатических градиентах. Выявленные закономерности изменения БП древесины и коры деревьев в градиентах температур и осадков полностью повторяют ранее установленные закономерности изменения фитомассы и чистой первичной продукции деревьев и древостоев Евразии в тех же градиентах. Это означает, что климатическая обусловленность биологической продуктивности имеет общий характер как для количественных, так и для квалиметрических показателей деревьев и древостоев
Предпросмотр статьи





Идентификаторы и классификаторы
- SCI
- Биология
В условиях изменяющегося климата возрастает биосферная роль лесного покрова и актуальность исследований углерододепонирующей способности мировых лесов. Сегодня формируются глобальные базы данных не только о фитомассе деревьев и древостоев (Falster et al., 2015; Kattge et al., 2020), но и о квалиметрических показателях фитомассы, в частности, базисной плотности (БП) стволовой древесины (Zanne et al., 2009; Usoltsev, 2020), составляющей основную часть надземной фитомассы. Эти базы данных предоставляют необходимую исходную информацию для глобальных прогнозов углерододепонирующей способности лесов. Показатель плотности древесины, используемый при преобразовании данных о запасе стволовой древесины в показатели фитомассы, является ключевым фактором, влияющим на точность оценки депонирования углерода (Fearnside, 1997; Усольцев, Цепордей, 2020). С другой стороны, изменения климата, в частности учащающиеся засухи в некоторых регионах, влияют на физиологические процессы, определяющие ксилогенез и БП древесины (Bouriaud et al., 2005; Vieira et al., 2020)
Список литературы
1. Андреева И.И., Родман Л.С. Ботаника. М.: КолосС, 2003. 528 с.
2. Дрейпер Н., Смит Г. Прикладной регрессионный анализ. М.: Статистика, 1973. 392 с.
3. Лиепа И.Я. Динамика древесных запасов: Прогнозирование и экология. Рига: Зинатне, 1980. 170 с.
4. Полубояринов O.И. Плотность древесины. М.: Лесн. пром-сть, 1976. 160 с.
5. Исаева Л.Н. Метод расчета локальной и средней плотности абсолютно сухой древесины в стволах сосны и лиственницы // Лесоведение. 1978. № 4. С. 90-94.
6. Мелехов В.И., Бабич Н.А., Корчагов С.А. Качество древесины сосны в культурах. Архангельск: Изд-во АГТУ, 2003. 110 с.
7. Усольцев В.А., Цепордей И.С. Квалиметрия фитомассы лесных деревьев: плотность и содержание сухого вещества: монография. Екатеринбург: Уральский государственный лесотехнический университет, Ботанический сад УрО РАН, 2020. 182 с. (CD-ROM). https://elar.usfeu.ru/bitstream/123456789/10022/1/Usolcev_20.pdf. EDN: CFOFQO
8. Усольцев В.А., Цепордей И.С. Климатически обусловленные территориальные изменения фитомассы деревьев лесообразующих видов Евразии и их прогнозирование // Сибирский лесной журн. 2021а. № 6. С. 72–90. EDN: TNPLOD
9. Усольцев В.А., Цепордей И.С. Принцип пространственно-временнóго замещения в экологии и прогнозирование биомассы Picea spp. при изменении климата // Хвойные бореальной зоны. 2021б. Т. 39. № 4. С. 269–275. EDN: ZICLFK
10. Усольцев В.А., Цепордей И.С., Азаренок М.В. Климатически обусловленные пространственные и темпоральные изменения биомассы рода Abies spp. Евразии в контексте закона лимитирующего фактора // Хвойные бореальной зоны. 2021. Т. 39. № 5. С. 392–400. EDN: IBTJFM
11. Фонти М.В. Климатический сигнал в параметрах годичных колец (плотности древесины, анатомической структуре и изотопном составе) хвойных и лиственных видов деревьев в различных природно-климатических зонах Евразии: дис. … докт. биол. наук: 03.02.08. Красноярск: СибФУ, 2020. 45 с.
12. Baskerville G.L. Use of logarithmic regression in the estimation of plant biomass // Canadian J. Forest Research. 1972. V. 2. P. 49-53.
13. Billard A., Bauer R., Mothe F., Colin F., Longuetaud F. Vertical variations in wood basic density for two softwood species // European J. Forest Research. 2021. V. 140. P. 1401–1416. EDN: PNSDEV
14. Bouriaud O., Leban J.-M., Bert D., Deleuze C. Intra-annual variations in climate influence growth and wood density of Norway spruce // Tree Physiology. 2005. V. 25. P. 651–660. EDN: XSUKKY
15. Castro V.R., Chambi-Legoas R., Filho M.T., Surdi P.G., Zanuncio J.C., Zanuncio A.J.V. The effect of soil nutrients and moisture during ontogeny on apparent wood density of Eucalyptus grandis // Scientific Reports. 2021. V. 11. № 1. Article 2530. DOI: 10.1038/s41598-020-59559-2 EDN: VGPEAZ
16. Falster D.S., Duursma R.A., Ishihara M.I., Barneche D.R., FitzJohn R.G., Vårhammar A., Aiba M., Ando M., Anten N., Aspinwall M.J., Baltzer J.L., Baraloto C., Battaglia M., Battles J.J., Bond-Lamberty B., van Breugel M., Camac J., Claveau Y., Coll L., Dannoura M., Delagrange S., Domec J.-C., Fatemi F., Feng W., Gargaglione V., Goto Y., Hagihara A., Hall J.S., Hamilton S., Harja D., Hiura T., Holdaway R., Hutley L.S., Ichie T., Jokela E.J., Kantola A., Kelly J.W.G., Kenzo T., King D., Kloeppel B.D., Kohyama T., Komiyama A., Laclau J.-P., Lusk C.H., Maguire D.A., le Maire G., Mäkelä A., Markesteijn L., Marshall J., McCulloh K., Miyata I., Mokany K., Mori S., Myster R.W., Nagano M., Naidu S.L., Nouvellon Y., O’Grady A.P., O’Hara K.L., Ohtsuka T., Osada N., Osunkoya O.O., Peri P.L., Petritan A.M., Poorter L., Portsmuth A., Potvin C., Ransijn J., Reid D., Ribeiro S.C., Roberts S.D., Rodríguez R., Saldaña-Acosta A., Santa-Regina I., Sasa K., Selaya N.G., Sillett S.C., Sterck F., Takagi K., Tange T., Tanouchi H., Tissue D., Umehara T., Utsugi H., Vadeboncoeur M.A., Valladares F., Vanninen P., Wang J.R., Wenk E., Williams R., Ximenes F. de Aquino, Yamaba A., Yamada T., Yamakura T., Yanai R.D., York R.A. BAAD: a Biomass And Allometry Database for woody plants // Ecology. 2015. V. 96. Article 1445.
17. Fearnside P.M. Wood density for estimating forest biomass in Brazilian Amazonia // Forest Ecology and Management. 1997. V. 90. № 1. P. 59–89. EDN: AIYVKV
18. Freese F. Linear regression methods for forest research. USDA Forest Service. Res. Paper FPL 17. Madison, 1964. 136 p.
19. Fu L.Y., Zeng W.S., Tang S.Z., Sharma R.P., Li H.K. Using linear mixed model and dummy variable model approaches to construct compatible single-tree biomass equations at different scales – A case study for Masson pine in Southern China // J. Forest Science. 2012. V. 58. № 3. P. 101–115. EDN: PNYGTH
20. Fujimoto T., Kita K., Kuromaru M. Genetic control of intra-ring wood density variation in hybrid larch ( Larix gmelinii var. japonica × L. kaempferi ) // Wood Science and Technology. 2008. V. 42. P. 227–240.
21. Howe J.P. Relationship of climate to the specific gravity of four Costa Rican hardwoods, an exploratory study // Wood Fiber. 1974. V. 5. P. 347–352.
22. Jacobs M.W., Cunia T. Use of dummy variables to harmonize tree biomass tables // Canadian J. Forest Research. 1980. V. 10. № 4. P. 483–490.
23. Kattge J., Bönisch G., Díaz S., Lavorel S., Prentice I.C., Leadley P., Tautenhahn S. et al. TRY plant trait database – Enhanced coverage and open access // Global Change Biology. 2020. V. 26. P. 119–188. EDN: BXAOPJ
24. Kellomäki S. On geoclimatic variation in basic density of Scots pine wood // Silva Fennica. 1979. V. 13. № 1. P. 55–64.
25. Kerfriden B., Bontemps J.-D., Leban J.-M. Variations in temperate forest stem biomass ratio along three environmental gradients are dominated by interspecific differences in wood density // Plant Ecology. 2021. V. 222. № 3. P. 289–303. EDN: LFHNOY
26. Lachowicz H., Bieniasz A., Wojtan R. Variability in the basic density of silver birch wood in Poland // Silva Fennica. 2019. V. 53. № 1. Article id 9968.
27. Mikola P. On variations in tree growth and their significance to growth studies // Communicationes Instituti forestalis Fenniae. 1950. V. 38. № 5. P. 1–131.
28. Molteberg D., Høibø O. Modelling of wood density and fibre dimensions in mature Norway spruce // Canadian J. Forest Research. 2007. V. 37. № 8. P. 1373–1389.
29. Saikku O. The effect of fertilization on the basic density of Scots pine ( Pinus silvestris L.). A densitometric study on the X ray chart curves of wood // Communicationes Instituti forestalis Fenniae. 1975. V. 85. № 3. P. 1-49.
30. Sousa V.B., Louzada J.L., Pereira H. Age trends and within-site effects in wood density and radial growth in Quercus faginea mature trees // Forest Systems. 2016. V. 25. № 1. Article e053.
31. St-Germain J.-L., Krause C. Latitudinal variation in tree-ring and wood cell characteristics of Picea mariana across the continuous boreal forest in Quebec // Canadian J. Forest Research. 2008. V. 38. № 6. P. 1397–1405. EDN: MJDJXN
32. Swenson N.G., Enquist B.J. Ecological and evolutionary determinants of a key plant functional trait: wood density and its community – wide variation across latitude and elevation // American J. Botany. 2007. V. 94. № 3. P. 451–459. EDN: KGYOFU
33. Télles J.R.G., Martínez A.V., de la Rosa A.B., Grande J.C., Mendoza C.P. Radial variation of basic density in Pinus patula Schltdl. et Cham. in three locations from Hidalgo state // Revista Mexicana de Ciencias Forestales. 2011. V. 2. № 7. P. 71–78.
34. Usoltsev V.A. Stem taper, density and dry matter content in biomass of trees growing in Central Eurasia: CD-monograph. Yekaterinburg: Ural State Forest Engineering University, Botanical Garden of Ural Branch of RAS, 2020. Available at: https://elar.usfeu.ru/handle/123456789/9649.
35. Usoltsev V.A., Shobairi S.O.R., Tsepordey I.S., Chasovskikh V.P. On some differences in the response of Picea spp. and Abies spp. single-tree biomass structure to changes in temperatures and precipitation in Eurasia // Environment and Ecology. 2020a. V. 38. P. 300–315. EDN: KUVLPH
36. Usoltsev V.A., Shobairi S.O.R., Tsepordey I.S., Chasovskikh V.P. Additive model of aboveground biomass of larch single-trees related to age, DBH and height, sensitive to temperature and precipitation in Eurasia // J. Applied Sciences & Environmental Management. 2020b. V. 24. P. 1759–1766.
37. Usoltsev V., Zukow W., Tsepordey I. Climatically determined spatial and temporal changes in the biomass of Pinus sp. of Eurasia in the context of the law of the limiting factor // Ecological Questions. 2022. V. 33. № 1. P. 15–23. DOI: 10.12775/EQ.2022.007 EDN: CLMNSI
38. Vieira J., Carvalho A., Campelo F. Tree growth under climate change: Evidence from xylogenesis timings and kinetics // Frontiers in Plant Science. 2020. V. 11. Article 90. EDN: BMJTNV
39. Wiemann M.C., Williamson G.B. Geographic variation in wood specific gravity: effects of latitude, temperature and precipitation // Wood and Fiber Science. 2002. V. 34. № 1. P. 96–107.
40. World Weather Maps, 2007. https://www.mapsofworld.com/ referrals/weather.
41. Zanne A.E., Lopez-Gonzalez G., Coomes D.A., Ilic J., Jansen S., Lewis S.L., Miller R.B., Swenson N.G., Wiemann M.C., Chave J. Global wood density database. Dryad, 2009. Identifier: http://hdl.handle.net/10255/dryad.235.
42. Zeng W.S. Developing tree biomass models for eight major tree species in China. In: Biomass volume estimation and valorization for energy. Chapter 1. Intech Publ., 2017. P. 3–21.
43. Zhang L., Shi H. Local modeling of tree growth by geographically weighted regression // Forest Science. 2003. V. 50. № 2. P. 225–244.
Выпуск

Другие статьи выпуска

В статье приведен анализ исторических данных по организации и состоянию геоэкологических исследований в горных лесах России и Северного Кавказа. На примере регионального мониторинга горных лесных экосистем показана организация многоцелевого природопользования в пределах горных водосборных бассейнов. По результатам экспериментального материала, полученного многолетним мониторингом на комплексных лесогидрологических стационарах с постановкой активных экспериментов, даны рекомендации по лесопользованию в горах. Показан пример получения информации экспедиционными методами, а также компьютерное моделирование лесохозяйственного освоения в условиях горных регионов. Предложены научные принципы и экологические основы ведения хозяйства в лесах горных регионов страны

Опасность массовых размножений стволовых вредителей в Ленинградской области и Республике Карелии возрастает. Однако данные по состоянию их популяций на северо-западе европейской части России немногочисленны и весьма разнородны. Они включают материалы статей и обзоров санитарного и лесопатологического состояния, которые, в свою очередь, основываются на совершенно разных методических подходах к получению информации. Обобщение и анализ данных по увеличению численности стволовых вредителей, представляющих опасность для хвойных древостоев северо-запада европейской части России, с учетом материалов научных отчетов и лесопатологического мониторинга – актуальная задача данной работы. Наибольшую опасность для еловых древостоев представляет короед-типограф Ips typographus (Linnaeus, 1758), формирующий масштабные очаги массового размножения. Основные факторы, обуславливающие начало формирования очагов размножения, – погодные условия (ураганные ветры и образование ветровалов, увеличение температуры в течение вегетационного сезона и недостаток осадков). В конце XX–начале XXI в. частоты вспышек массовых размножений стволовых вредителей, в особенности короеда-типографа, в Ленинградской области увеличились. Начиная со второй половины XX в., отмечаются вспышки размножения в Республике Карелии. В Мурманской области стволовые вредители не оказывают заметного влияния на состояние древостоев. Эти тенденции соответствуют тренду увеличения температуры в Ленинградской области и Карелии и несущественному изменению температуры в Мурманской области. Дополнительное питание сосновых лубоедов и черных усачей в случаях их массового размножения – недооцененный фактор ослабления древостоев, который существенно влияет на прирост и состояние окружающих насаждений. Появление инвазивных видов, таких как союзный короед Ips amitinus (Eichhoff, 1872) (Curculionidae: Coleoptera), представляет потенциальную опасность, однако в настоящее время в сосновых и еловых лесах на севере европейской части России этот вид не проявляет существенной активности

Изучались торфянистые подстилки мохово-древесного состава в насаждениях березы пушистой (Betula pubescens Ehrh.) травяно-мшистых групп типов леса в северной части междуречья Оби и Томи (географические координаты 56°23′186′′ с. ш., 084°32′519′′ в. д.). Подстилки характеризуются слабой насыщенностью основаниями – 35.6%, кислой реакцией среды – 4.0, зольностью – 8.9%. Среднее содержание макро- и микроэлементов образует следующий нисходящий ряд, мг/кг: Сa5105 > > Fe 4201 > Al3614 > K 986 > Mg893 > Mn 468 > Sr230 > Na 153 > Zn51 >Pb13 > Cu6.8 > Ni5 > Cr4.6 > Co 2.9 > Cd0.2. Согласно факторному анализу, состав минеральной компоненты подстилок в процессе деструкции растительных остатков на 57% определяется изменчивостью содержания тяжёлых металлов, на 37% – щелочных, щелочноземельных, Zn, Cd. Методом древовидной кластеризации морфометрические фракции растительных фрагментов (дериваты) организовались следующим образом: >10 мм, [(10–5) + + (5–3) + (3–2) + (2–1)] и [(1–0.5) + (0.5–0.25) + <0.25] мм. Дискриминантный анализ показал 100% попадание в соответствующую группу. Наибольший вклад в предсказание вносят Al и Са. По мере распада растительных фрагментов тяжелые металлы и алюминий последовательно накапливаются, щелочные и щелочноземельные металлы вымываются и наиболее активно – на стадии ферментации. Распределение минеральных компонентов в дериватах подстилок согласуется с их гумусным состоянием. В мелких морфометрических фракциях по сравнению с крупными фрагментами расширяется отношение (∑ГК + ∑ФК)/полисахариды, сужается величина С/N, накапливаются гуминовые и фульвокислоты главным образом 1-й фракции. Торфянистые подстилки характеризуются средним, умеренно опасным уровнем загрязнения: суммарный показатель загрязнения (Zc) – 18. В составе токсикантов доминируют Pb, Zn – химические элементы I класса опасности

В статье обсуждаются результаты комплексного анализа экологического состояния и средостабилизирующих функций видового состава арборифлоры в структуре городского озеленения. В основу работы положен большой фактический материал, полученный в ходе многолетнего мониторинга зеленых насаждений г. Владивостока. Разработано методическое обеспечение качественно-количественной оценки видов с использованием методов прикладной квалиметрии. В качестве единицы оценки предложен интегральный показатель – коэффициент функциональной эффективности вида (КФЭВ) в озеленении. Он представляет собой относительно-количественный показатель качества, определяемый по совокупности ряда эколого-биологических и санитарно-гигиенических свойств растений: распространенности в озеленении, жизненного статуса, способности к аккумуляции приоритетных металлов-загрязнителей в городской среде, их концентрации относительно локального экологического фона, интенсивности накопления металлов из почвы. На основе предложенного коэффициента выполнен сравнительный анализ функциональной эффективности 80 видов деревьев и кустарников, формирующих городские насаждения Владивостока. В сравниваемой выборке растений КФЭВ снижается от 3.70 у боярышника перистонадрезанного (Crataegus pinnatifida) до 1.13 у яблони маньчжурской (Malus mandshurica). Эти показатели соответствуют 74 и 23% принятого стандарта качества (СК) видов. Выделены группы видов разной функциональной значимости в городском озеленении. Наибольшую эффективность в создании комфортных для проживания горожан условий среды показали виды широкого распространения в зеленых насаждениях: ясень маньчжурский (Fraxinus mandshurica), вяз японский (Ulmus japonica), берёза плосколистная (Betula platyphylla), пузыреплодник калинолистный (Physocarpus opulifolia) и др. Для них характерно как максимальное участие в формировании структуры городского озеленения, так и высокая способность к поглощению основных металлов-загрязнителей городской среды. КФЭВ этих видов находится в пределах 3.26–2.61, что соответствует 65–52% СК. В заключении даны предложения по рациональному использованию видов в формировании комфортной городской среды и внедрению полученных результатов в практику управления городским зеленым фондом

Приведены данные по влиянию колоний серой цапли (Ardea cinerea L., 1758) на динамику параметров годичного кольца деревьев в сосняках искусственного происхождения. Показано, что начало колонизации насаждений серой цаплей можно довольно точно установить при анализе динамики параметров годичных колец деревьев, особенно ширины их позднего слоя и оптической плотности древесины (значения первого из этих параметров в контрольном насаждении ниже, а второго выше). Колонии серой цапли начинают появляться в сосняках уже с 10–12-летнего возраста, когда высота деревьев достигает 5–6 м, а диаметр ствола на высоте 1.3 м от поверхности почвы – всего 7–8 см. В первые 15–20 лет они оказывают положительное влияние на годичный прирост деревьев, особенно на ширину позднего слоя древесины и толщину клеточных стенок, но затем картина меняется на противоположную. Общее время пребывания колоний цапли на одном месте составляет около 35– 40 лет, после чего они переселяются на новые места в связи с расстройством или же полной гибелью насаждений. Для снижения вероятности появления колоний серой цапли, которые во многих случаях оказывают отрицательное влияние на состояние лесных биогеоценозов, необходимо отказаться от создания вблизи водоемов чистых сосняков, отдавая предпочтение еловым, березовым, тополевым или же липово-дубовым насаждениям

Изучены культуры 1Б класса бонитета полнотой 0.95 на площади 0.64 га, созданные на вырубке по схеме 1.82 × 1.10 м. На план наносили живые и отпавшие деревья и площади их питания в программе “ArcMap-ArcView”. Территорию разделили на 9 секций, с густотой 76–122% от среднего по насаждению. Доставшаяся деревьям в возрасте 30 лет площадь питания повлияла на диаметр ствола в 55 лет в редких местах древостоя с силой 13.3%, а в густых местах – с силой 5.0%. То есть подтвердилась гипотеза, что влияние площади питания дерева на диаметр ствола может быть слабым также и в среднем возрасте насаждений, и уровень его влияния зависит от густоты древостоя. Отпад деревьев к 55 годам коррелировал с частотой в классах площади питания (r = 0.96 ± 0.03), поэтому площадь питания менее среднего значения повышала вероятность отпада дерева к 55 годам всего лишь на 7%. Выдвинуто предположение, что снижение влияния площади питания при высокой густоте происходит из-за усиления кооперации деревьев. При имитационном разреживании культур, с увеличением площади питания у оставляемых деревьев в 2 раза было получено соответствующее увеличение диаметра только у 11% деревьев. Остальные 89% деревьев не воспользовались доставшейся им большей площадью питания и не увеличили свои размеры, несмотря на 25 лет развития при более свободном стоянии. Это указывает на то, что в культурах второго класса возраста увеличение площади питания деревьев уже не приводит к улучшению их развития в подавляющем числе случаев. Поэтому густоту следует снижать в намного более раннем возрасте, например, в 10–15 лет

Обсуждаются проблемы перехода сосны обыкновенной (Pinus sylvestris L.) из состояния устойчивого к неустойчивому равновесию, смены равновесной популяции на неравновесную в результате потепления климата. Цель исследований – изучить состояние генеративной сферы южной и центральной лесостепных популяций сосны обыкновенной; по признаку полнозернистости оценить их жизненное состояние в 2020 г. по сравнению с 2016 – годом дестабилизации сосновых лесов, а также относительно состояния степной популяции в урожайном 2017 г. Объектами исследования служили центральная и южная лесостепные популяции сосны, произрастающие на экологически благоприятной территории Воронежской и Белгородской областей. В 2015 г. зафиксирован переход их из равновесия в слабо неравновесную систему. Уровень полнозернистости ступинской и белгородской популяций снизился соответственно на 25.6 и 24.6%, число семян – на 37.7 и 22.4%, уровень смертности семяпочек повысился в 3.9 и 4.0 раза. Сравнение показателей семенной продуктивности в 2013, 2016 и 2020 гг. показало, что в настоящее время оба насаждения представляют равновесные системы. Центральная популяция вернулась к региональной норме (устойчивого равновесия), южная находится в состоянии неустойчивого равновесия. Дисперсионный анализ выявил существенное (57.8%) влияние фактора “ГТК” на генеративную сферу белгородской популяции по признаку полнозернистости. В последнее десятилетие число оптимальных лет в ЦЧР уменьшилось с 7–8/10 лет до 5. Для возвращения сосновых лесов в равновесие требуется 3 оптимальных года. Высокие темпы потепления климата могут повлечь за собой нарушение равновесия и повторную дестабилизацию. Дальнейшее сокращение числа оптимальных лет делает почти невозможным возвращение сосновых лесов к региональной норме

Изучение динамики парцеллярной структуры лесов Приморского края необходимо для выявления закономерностей естественного восстановления уникальной формации региона – коренных хвойно-широколиственных лесов. В результате хозяйственной деятельности эти леса, насыщенные реликтовыми элементами, сменились производными на большей части ареала. Исследования проводились в водосборном бассейне р. Комаровки (Южное Приморье, Уссурийский район). В статье приведены результаты изучения парцеллярной структуры широколиственно-липового с лианами лещинового разнотравного типа леса – репрезентативного для региона и самого сложного в формации производных лесов. Метод исследований классический: на постоянной пробной площади (ППП) выполнено детальное описание и картирование в масштабе 1: 100 всех ярусов фитоценоза с последующим попарным сравнением картосхем и выделением парцелл. Анализ трансформации фитоценоза за 20 лет восстановительной сукцессии показал, что основные лесоводственно-таксационные характеристики насаждения изменились незначительно. В древостое почти полностью выпала осина и значительная часть одной из главных пород – липы. Одновременно укрепились позиции подлеска и видов деревьев, характерных для коренных хвойно-широколиственных лесов. Изменения в парцеллярной структуре свидетельствуют об успешном восстановлении коренного типа леса: в фитоценозе число парцелл уменьшилось с 12 до 10; к шести условно-коренным добавилась еще одна. Общими остались 6 парцелл, все они условно-коренные. Произошло закономерное укрупнение и условно-коренных парцелл и выравнивание их границ. Отражена высокая индикаторная роль травяного яруса
Статистика статьи
Статистика просмотров за 2025 - 2026 год.
Издательство
- Регион
- Россия, Москва
- Почтовый адрес
- 117997, Российская Федерация, г. Москва, ул. Профсоюзная, 84/32, стр. 14
- Юр. адрес
- 117997, Российская Федерация, г. Москва, ул. Профсоюзная, 84/32, стр. 14
- ФИО
- Лукина Наталья Васильевна (Директор)
- E-mail адрес
- cepfras@cepl.rssi.ru
- Контактный телефон
- +7 (499) 7430016
- Сайт
- http:/cepl.rssi.ru